《Nature》子刊:破解肠脑轴迷走神经失调机制,靶向神经修复减重
原创
《Nature》子刊:破解肠脑轴迷走神经失调机制,靶向神经修复减重
原创
本草音乐实验室
发布于 2026-07-13 10:53:29
发布于 2026-07-13 10:53:29

当我们面对一块诱人的巧克力蛋糕或是一盘香气四溢的炸鸡时,为什么即使肚子已经感觉到了饱胀,大脑却依然不断发出“再吃最后一口”的强烈指令?
这并非单纯的“意志力薄弱”或“贪吃”,而是一场发生在我们的胃肠道与大脑中枢之间的精密信号战。在这场看不见的战役中,一根被称为“迷走神经”的超级信息高速公路,决定了我们何时渴望食物、何时停止咀嚼,以及最终的体型走向。
2026年4月21日,国际顶级学术期刊《Nature Reviews Gastroenterology & Hepatology》发表了一篇极具突破性的重磅综述文章。研究团队以前所未有的深度,系统剖析了“肠-脑轴”在饮食行为和肥胖发生中的核心调控作用。
这项研究不仅揭示了迷走神经作为人体内部“内感受”通讯光缆的精密运作机制,更首次系统性地提出了高脂高糖饮食诱导肥胖的“三阶段渐进演变模型”。
对于深受减重反弹困扰的社会公众而言,这项研究提供了一个极具温度且科学的解释;而对于临床医生和科研人员,它则为打破传统单一药物治疗的局限、开发下一代靶向神经修复的抗肥胖疗法,指明了全新的战略方向。
传统节食反弹与单一药物的现实困境
肥胖已经跨越了单纯的体型焦虑,演变为21世纪最具破坏性的全球公共卫生危机之一。根据世界卫生组织发布的最新权威流行病学数据,全球已有超过10亿人患有肥胖症,这一数字在过去三十年里呈现出爆炸式的指数级增长[1]。
肥胖不仅仅是皮下脂肪的堆积,它更是2型糖尿病、心血管疾病、非酒精性脂肪肝以及多种恶性肿瘤的直接导火索。这种沉重的慢性疾病负担,正在无情地压垮各国的医疗卫生系统,消耗着巨额的公共健康资源。
长期以来,我们对抗肥胖的主要武器被简单粗暴地归结为“管住嘴、迈开腿”。然而,海量的临床随访数据无情地揭示了一个残酷的真相:单纯依靠生活方式干预和节食的长期成功率极低。
大多数患者在经历痛苦的节食和剧烈的体重下降后,都会陷入一个令人绝望的死循环。体重的快速流失会触发身体的生存防御机制,导致基础代谢率断崖式降低,同时食欲激素疯狂反扑,最终引发体重的报复性反弹。
这种被称为“溜溜球效应”的减重模式,不仅一次次摧毁了患者的自信心,更让身体的内分泌和代谢系统在反复折腾中雪上加霜。临床上亟需一种能够真正打破这一恶性循环的破局之法。
近年来,以司美格鲁肽和替尔泊肽为代表的GLP-1受体激动剂横空出世,在临床试验中取得了堪比减重手术的奇迹般效果。然而,医学界很快发现了隐藏在奇迹背后的隐忧。
顶级医学期刊《新英格兰医学杂志》(NEJM)发表的STEP 1延伸试验表明,患者在停止注射司美格鲁肽一年后,此前减掉的体重会反弹超过三分之二,同时各项心血管代谢指标也随之恶化[2]。
这一现象暴露出一个严峻的临床未满足需求:现有的明星药物虽然能够通过模拟肠道激素强效抑制中枢食欲,但它们并未从根本上修复患者体内已经受损的“肠-脑通讯系统”。一旦外源性的药物干预撤除,失调的底层神经回路立刻会让肥胖卷土重来。
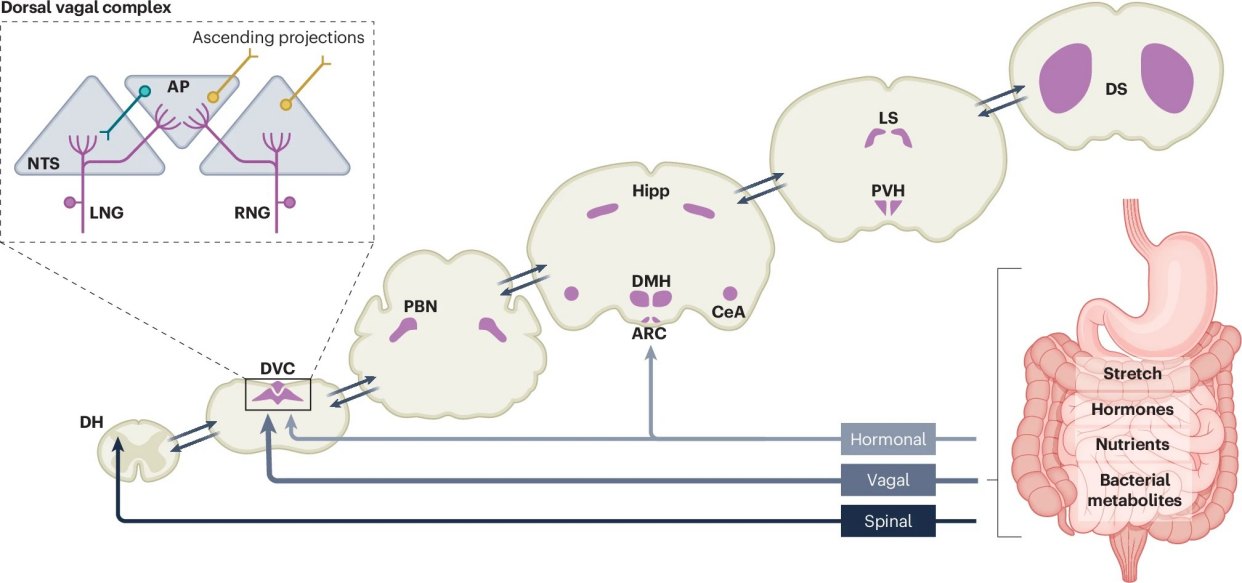
跨物种追踪与单细胞测序,精准解码神经元的多维技术矩阵
虽然本文是一篇高屋建瓴的权威综述,但其核心结论并非空中楼阁,而是建立在过去数十年海量、顶尖的原始实验数据之上。科学家们运用了一系列令人惊叹的现代现代神经科学技术,才得以窥见肠脑轴的微观世界。
研究整合了从果蝇、啮齿类动物(小鼠和大鼠)到人类的跨物种庞大数据库。在动物模型中,研究者摒弃了早期粗糙的“迷走神经切断术”,转而采用极其精密的现代回路追踪技术。
通过光遗传学和化学遗传学工具,科学家们能够特异性地标记并操控支配肠道的特定迷走神经感觉神经元。这使得研究人员可以在动物自由进食的状态下,实时观察开启或关闭某一条神经回路对食量的直接影响。
在人类研究层面,科学家则利用功能性磁共振成像和血氧水平依赖信号技术,实时捕捉受试者在摄入不同营养物质或看到食物图片时,大脑奖赏中枢(如纹状体、杏仁核)的动态激活过程。
为了精准拆解迷走神经内部极其复杂的功能分工,研究人员还引入了前沿的单细胞RNA测序技术。他们成功将迷走神经元划分为多个具有不同分子特征和专属职责的亚群。
例如,表达GLP1R和Oxtr基因的神经元主要分布在胃部肌肉层,负责感知胃壁的“物理拉伸”;而表达Cckar和Vip基因的神经元,则专门潜伏在肠道黏膜中,负责探测流经的“脂肪和糖分”。这种高分辨率的特征构建,为后续的机制解析奠定了坚实的数据基础。
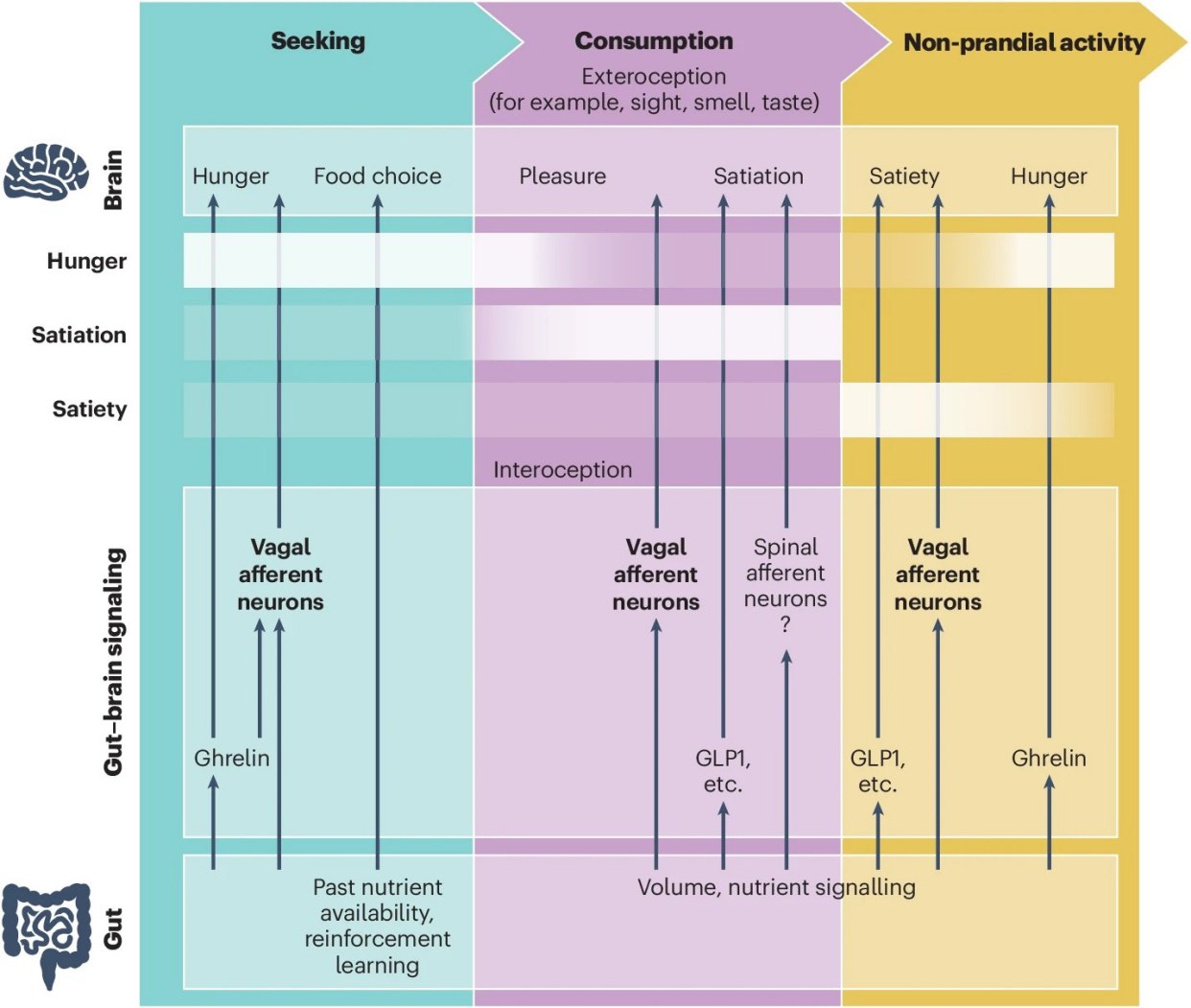
肠脑轴的“三相调控”机制,迷走神经如何全天候掌控食欲
在这项研究中,科学家将人类看似连续的饮食行为,极其精妙且科学地划分为三个独立的阶段:食物搜寻(觅食)、食物消耗(进食)以及非进食活动(饱腹期)。在这三个阶段的无缝切换中,迷走神经扮演了全天候的“最高情报官”。
觅食阶段:饥饿信号与多巴胺的狂热共舞
当我们的胃部空空如也,身体处于能量负平衡状态时,胃底的内分泌细胞会大量分泌一种被称为“饥饿素”的激素。这些信号不仅通过血液循环,更通过迷走神经上的GHSR受体,迅速将“电报”传导至大脑。
这些信号会精准激活下丘脑中的AgRP神经元,这正是控制我们食欲的“主开关”。一旦开关被按下,强烈的觅食动机便油然而生。
与此同时,肠道传来的饥饿信号会高度敏化中脑边缘和黑质纹状体的多巴胺系统。这使得我们在饥饿状态下,对食物的视觉、嗅觉等外部线索异常敏感。这完美解释了为什么在饥肠辘辘的下班路上,街边哪怕最普通的烤肉香气,也会显得具有致命的诱惑力。

进食阶段:物理拉伸与化学激素的双重刹车
当我们终于坐下开始大快朵颐,食物顺着食道进入胃肠道,迷走神经立刻从“催促者”变身为“监督员”。分布在胃壁肌层中的机械感受器(如IGLEs结构),会敏锐地感知到胃壁随着食物充盈而产生的物理拉伸。
这种物理拉伸信号,通过迷走神经末梢上的PIEZO等机械敏感离子通道,瞬间转化为高频电信号,沿着神经纤维飞速传导至脑干的孤束核(NTS)。这是身体踩下的第一脚“物理刹车”。
紧接着,当食物中的营养物质进入肠道,肠道黏膜上的内分泌细胞(EECs)在接触到脂肪、糖分和蛋白质后,会迅速释放GLP-1、CCK(胆囊收缩素)和PYY等化学激素。
这些化学信号不仅通过血液缓慢起效,更会直接作用于附近的迷走神经化学感受器。甚至有研究发现,肠道细胞会伸出被称为“神经足”(Neuropods)的结构,在亚秒级的时间内将谷氨酸直接释放给迷走神经[3]。物理与化学信号在脑干完美汇合,共同踩下死刹车,促使我们放下手中的刀叉。
饱腹阶段:长效的能量守卫与记忆烙印
当我们停止进食,进入漫长的非进食期,肠脑轴的工作并未停止。此时,肠道仍在持续消化吸收,迷走神经会持续向大脑发送“营养储备充足”的维稳信号。
这些信号会持续抑制下丘脑的饥饿神经元,从而有效延长两餐之间的间隔时间,防止我们刚吃完饭又想吃零食。
更有趣的是,迷走神经还会将这一餐的营养价值和愉悦体验,精准地传递给大脑的海马体,形成深刻的“食物记忆”。这种带有营养标签的记忆烙印,将潜移默化地指导我们下一次的觅食选择,形成一个完美闭环。
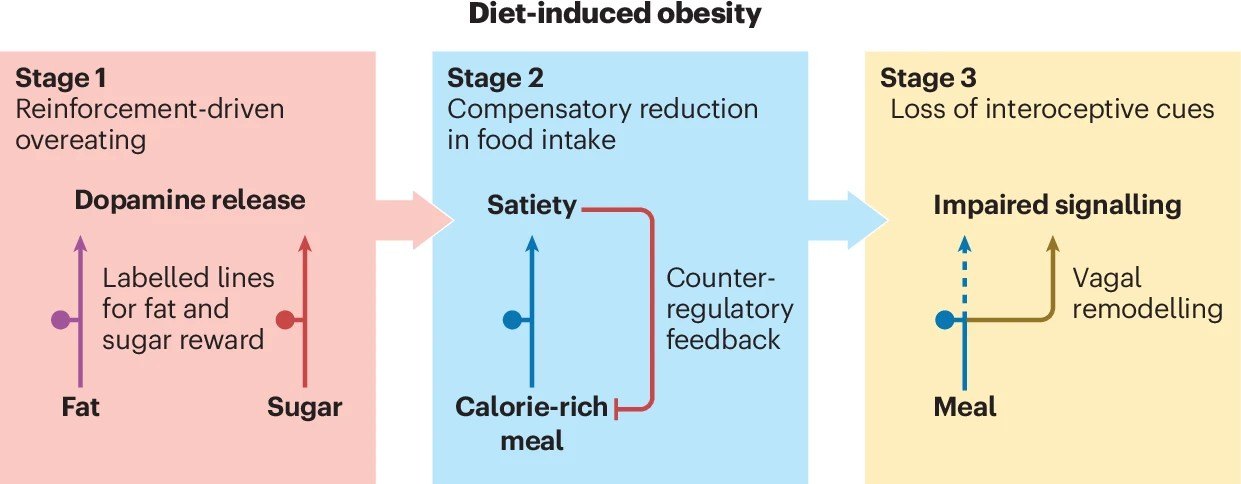
饮食诱导肥胖的三阶段演变,从多巴胺劫持到内感受信号全面崩盘
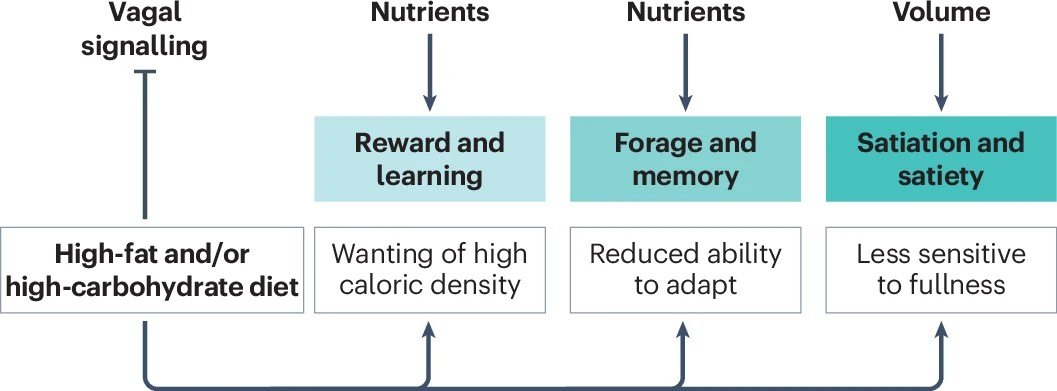
本研究最核心的理论突破,在于首次系统性地提出了长期摄入高脂高糖饮食(典型的现代西方饮食)摧毁肠脑轴的三个渐进阶段。这一演变模型,完美且残酷地解释了肥胖是如何一步步走向失控的。
阶段一:强化驱动的过度进食与多巴胺劫持
在接触高热量、高脂高糖食物的初期,我们的身体其实并未立刻发胖,但大脑的底层逻辑已经被悄然篡改。肠道内高浓度的脂肪和糖分,会极其强烈地激活迷走神经的化学感受器。
这种超强信号传导至大脑后,会引发纹状体释放海量的多巴胺。研究证实,当脂肪和糖分同时存在时(如冰淇淋、奶茶),它们对奖赏中枢的激活程度远超单一营养素,产生了一种“超加和效应”[4]。
这种强烈的正向反馈,让大脑迅速建立起“高热量=极度快乐”的牢固条件反射。此时,个体对高脂高糖食物产生了难以抗拒的偏好,觅食动机被极大地强化,为后续的肥胖埋下了第一颗定时炸弹。
阶段二:代偿性进食减少与身体的最后防线
面对持续的高热量轰炸,历经数百万年进化的人类躯体并未坐以待毙。在肥胖真正降临前,迷走神经会启动一系列强大的代偿机制,试图力挽狂澜。
其中最著名的机制被称为“回肠制动”(Ileal brake)。当大量未完全消化的超载营养物质到达远端小肠(回肠)时,肠道L细胞会大量释放GLP-1和PYY。这些激素通过迷走神经反射,迫使胃部肌肉放松,大幅减缓胃排空速度。
同时,进食释放的瘦素(Leptin)和CCK会产生协同作用,使得迷走神经对胃部的微小拉伸变得极其敏感。这使得动物或人类在摄入高热量食物时,会自觉地减少单次进食的物理体积,努力维持每天的总热量摄入不超标,从而在一段时间内死死守住体重的防线。
阶段三:内感受信号丧失与防线彻底崩溃
然而,如果高脂高糖饮食的恶习持续存在,迷走神经的代偿机制终将耗尽并走向衰竭。随着时间的推移,迷走神经对胃部拉伸和饱腹激素的敏感性出现了断崖式的下降。
这标志着肥胖演变进入了最具破坏性的第三阶段:内感受信号的全面丧失。此时,肠脑通讯彻底崩溃,胃肠道发出的“我已经吃撑了”的求救信号,根本无法传达到大脑。
更可怕的是,大脑下丘脑区域会产生严重的“瘦素抵抗”。尽管此时患者体内已经堆积了如山的脂肪,分泌了极高的瘦素,但大脑却“充耳不闻”,依然固执地认为身体正处于极度饥荒之中。于是,大脑下达了疯狂进食、囤积脂肪的最高指令,肥胖的深渊就此彻底敞开,再难回头。
迷走神经“断联”的微观真相,物理萎缩、菌群推波助澜与基因重写
为什么在第三阶段,原本尽职尽责的迷走神经会彻底罢工?科学家们深入微观的细胞与分子层面,揭示了令人震惊且不寒而栗的生物学机制。
突触末梢的实质性物理萎缩
研究表明,肠脑轴的断联并非仅仅是功能性的迟钝,而是发生了真实的物理结构损毁。在动物实验中,持续喂食高脂饮食仅仅3周后,小鼠脑干(NTS区域)中迷走神经的末梢密度就出现了显著的萎缩和减少。
这就好比连接肠道和大脑的“跨海光缆”发生了大面积的物理断裂。由于硬件的直接损坏,胃肠道发出的饱腹电信号在传输途中大量丢失,大脑接收到的永远是微弱的残缺信息,自然无法下达停止进食的指令。
肠道微生态失调的推波助澜
这种神经萎缩并非无缘无故。外部的独立研究早已证实,高脂高糖饮食会迅速且剧烈地改变肠道微生态,导致厚壁菌门(Firmicutes)急剧扩张,而拟杆菌门(Bacteroidetes)大幅减少[5]。
这种失衡的有害菌群会产生大量的促炎代谢产物和内毒素。这些毒素穿透受损的肠道屏障,直接攻击黏膜下的迷走神经末梢,引发持续的局部神经炎症。
令人惊叹的是,实验证明,如果将肥胖小鼠的粪便菌群提取出来,移植给原本健康的瘦小鼠,瘦小鼠的迷走神经也会迅速出现同样的萎缩现象,并开始暴饮暴食。这铁证如山地表明,肠道菌群是摧毁迷走神经的重要帮凶。
神经元基因表达图谱的全面重写
在长期的炎症浸润和高胰岛素、高瘦素的恶劣微环境中,迷走神经元内部的基因表达图谱发生了根本性的重编程。
原本负责传递饱腹感的关键受体(如机械敏感离子通道和CCK受体)的基因表达量骤减。同时,一种名为CART(可卡因和安非他明调节转录物)的饱腹相关神经肽的表达也被严重抑制。
这种分子层面的彻底重写,使得迷走神经即使在面对海量的食物填塞时,也变得极度麻木和迟钝。患者因此彻底丧失了对进食量的自我感知能力,陷入了吃得越多、饿得越快的绝望境地。
从“治标”走向“治本”,靶向神经修复的下一代抗肥胖疗法与生活启示
这项发表在《Nature》子刊上的宏大综述,不仅是一次基础科学的辉煌胜利,更为未来亿万肥胖患者的临床管理,描绘了一幅充满希望的全新蓝图。
当前的GLP-1受体激动剂虽然光芒万丈,但它们主要作用于迷走神经下游的中枢神经回路,通过高浓度的药物强行压制食欲。它们强效且直接,但正如文章指出的,它们并未真正修复已经受损萎缩的迷走神经本身。
这完美解释了为什么一旦停药,患者极易面临报复性反弹。未来的临床策略,应当发生范式转变:将GLP-1类药物视为“启动减重的破冰船”,利用其创造的体重下降和代谢改善的宝贵窗口期,去实施更深层次的肠脑轴神经修复干预。
真正的治本之策,在于恢复肠脑轴通讯的保真度与敏感性。未来的前沿治疗方向将呈现出令人振奋的多样化趋势。
微生态靶向治疗: 通过精准的益生元干预或粪菌移植(FMT)技术,我们可以从源头上重塑患者的肠道微生态环境。清理有害菌群,消除肠道局部的神经炎症,从而为迷走神经末梢的重新生长和功能恢复提供肥沃的土壤。
无创神经调控: 利用经皮迷走神经刺激(tVNS)等先进的物理医疗设备,在耳甲或颈部进行温和的电刺激。这种方法可以人为地增强迷走神经的电生理活性,帮助那些已经麻木的神经元重新焕发活力,进而重新唤醒大脑深处的饱腹感中枢。
模拟减重手术的机制: 临床观察中有一个极其神奇的现象,接受胃旁路等减重手术的患者,不仅胃容积变小了,其迷走神经的敏感性也在术后奇迹般地迅速恢复。科学家们正致力于破解这一手术背后的分子机制,一旦成功,我们极有望开发出能够模拟手术效果的“口服神经修复药物”。
除了前沿的医疗干预,这项研究也为普通公众的日常健康管理提供了极其温暖且实用的启示。它告诉我们:肥胖往往不是因为你缺乏意志力,而是你的神经回路暂时“生病”了。
在日常生活中,我们可以通过顺应人体的生物钟来主动保护我们的肠脑轴。研究明确证实,迷走神经的敏感性具有极其明显的昼夜节律特征,白天敏锐,夜晚迟钝。
因此,将主要的进食时间限制在白天的活跃期(如采用轻断食或限时进食策略),并坚决避免在深夜摄入高脂高糖的夜宵,能够有效维持迷走神经的敏锐度,防止其因过度疲劳而陷入“迟钝”状态。
当然,目前的机制解析仍有其局限性,大量数据依然依赖于啮齿类动物模型。人类的饮食行为远比小鼠复杂得多,其中交织着深厚的社会文化背景、复杂的情感压力以及多态的遗传基因。
未来,医学界需要开展更多大规模、多中心的人类前瞻性队列研究。结合多模态脑成像技术、连续血糖监测以及可穿戴代谢设备,进一步验证这些精妙机制在人体中的确切表现。
但无论如何,科学的巨轮已经转向。将目光从单纯的“卡路里计算”与“意志力对抗”,转向“肠脑轴信号的精准修复”,必将引领下一代抗肥胖医学的伟大变革,让更多人重获健康与自由。
参考资料
[1]World Health Organization. (2024). Obesity and overweight Fact sheet.
[2]Wilding, J. P. H., Batterham, R. L., Davies, M., et al. (2022). Weight regain and cardiometabolic effects after withdrawal of semaglutide: the STEP 1 trial extension. Diabetes, Obesity and Metabolism, 24(8), 1553-1564.
[3]Kaelberer, M. M., Buchanan, K. L., Klein, M. E., et al. (2018). A gut-brain neural circuit for nutrient sensory transduction. Science, 361(6408), eaat5236.
[4]DiFeliceantonio, A. G., Coppin, G., Rigoux, L., et al. (2018). Supra-additive effects of combining fat and carbohydrate on food reward. Cell Metabolism, 28(1), 33-44.e3.
[5]Ley, R. E., Turnbaugh, P. J., Klein, S., & Gordon, J. I. (2006). Human gut microbes associated with obesity. Nature, 444(7122), 1022-1023.
[6]de Lartigue, G., Brierley, D. I., & Choi, H. J. (2026). The critical role of gut–brain signalling in eating behaviour and obesity. Nature Reviews Gastroenterology & Hepatology.
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
原创声明:本文系作者授权腾讯云开发者社区发表,未经许可,不得转载。
如有侵权,请联系 cloudcommunity@tencent.com 删除。
评论
登录后参与评论
推荐阅读
目录
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号